Dove Behavior, Sexing Doves I, Sexing Doves II, Sexing Doves III, Sexing Doves IV, Sexing 2-egg clutches
THE SEXES IN 2-EGG
CLUTCHES OF RINGNECKS,
Streptopelia risoria.
by
Wilmer
J. Miller and Paul Hollander
A myth concerning the sexes in the 2-egg clutches of doves and pigeons never has been erased completely. In W. M. Levi’s 1957 book The Pigeon, he covers the topic well for the domestic pigeon, Columba livia. Aristotle before 322 B.C. wrote "The pigeon as a rule, lays a male egg and a female egg and generally allows a day’s interval to ensue and then lays the second egg." Aelian (about 222 A.D.) is more extreme. "The female brings forth two eggs, the first of which always produces a male, the second a female."
Later authorities follow in a similar manner, although not quite so absolute, as can be noted in Fulton and Lumley (1895) "The two eggs which a pigeon lays are almost invariably cock and hen, the cock being almost invariably the first one".
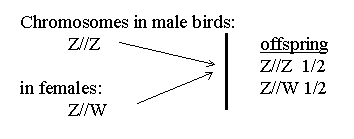
Levi after quoting such authorities correctly indicates the expected clutch ratio of 25% males, 50% males/females, and 25% females. This follows from the statistical knowledge of the laws of sex chromosomal segregation. In most species of birds the actual presence of the W is not proven and W could represent the absence of a chromosome leading to the same generalizations anyway. The slash represents the chromosome and is often omitted for brevity. One chromosome comes from the father and the other from the mother. Likewise one is transmitted to the offspring from the father and one from the mother. ZZ controls production of a male and ZW a female in birds (interacting with many other genes of course).
Of course, other factors might influence the ratio, but those known or suspected in organisms in general are minor. And why not actually look to see what it really is?!
Levi cites several such experiments including some tabulations of the Palmetto Pigeon Plant data by Dr. W. F. Hollander for sex-linked matings in the pigeon. Small deviations from the exact expected are expected, of course. But they are all within simple chance deviations.
With numbers in the thousands Levi slays the dragon of this myth.
But what about ringnecks, Streptopelia risoria, which is another genus and species. Some people on the internet have wondered about the sex distribution in the clutches. So I have started compiling the results of over 20 years of results of genetic breeding of ringnecks for other purposes.
(That number could be expanded to 60 years if small numbers of mating were included.)
I start here with results from the Iowa State University dove colony of 200 matings of birds descended from albino ringnecks imported from Japan. These doves were originally designated Streptopelia douraca. But they are placed within risoria now. And they are completely fertile with our familiar ringnecks. In fact the original birds in the importation of 3 males and 3 females were so infertile (from inbreeding?) that outcrosses had to be made to save the albino gene. Thus this population is more regular risoria than the original stock.
One bias in the study is that males evince their sex by bow-cooing at variable times but weeks to months before the females are evident by laying. Since many of the offspring had to be disposed of (usually donated), before being sexed, more males were known than females. Also keeping BOTH members of a clutch was infrequent, so that the numbers from this group are small.
Fortunately, ringnecks also have sex-linked colors allowing sex to be noted at hatching from particular appropriate matings. The sex-linked alleles controlling the dilution colors blond and white and the wild type dark allow the sex to be determined at hatching if a lighter colored male has a darker colored mate. The genes controlling these are labeled D+ for dark > dB for blond and dw for white in order of decreasing dominance.
Some other matings can be termed sex-linked partial matings, since half the daughters can be distinguished at hatching. For example the mating of dBdw x dB is one of this latter type. Half the daughters will be white and therefore female. The other half of the daughters will be blond and not distinguishable from blond males. So the results are divided into sex-linked matings, sex-linked partial and non-sex-linked (autosomal) matings.
The other methods of sexing were behavior (bow-coo from males); the "look-see" method using an infant nasal speculum inserted into the cloaca of mature individuals (Miller and Wagner ); and the actual mating success in producing offspring.
Since records were taken 3 times a week, and with experience in noting the squab’s development including the segregating color differences available, the first squab versus the second was almost always known. Color differences were usually evident at hatching so that the older squab could almost always be detected. Segregation and assortment were the general situation since genetic numbers to fit Mendelian ratios were being studied as well as blood types and other characters such as silky (Miller 1958?). In the minority of cases without color differences evident still the older squab could almost always be detected since the growth of the feathers filling in the area about the corner of the mouth was more advanced in the older squab. So Aelian’s notion that the first egg was the male could also be checked.
In doves the triple allelic series of sex-linked alleles are dark, blond, white in order of decreasing dominance. For example, a dark female mated to a blond male resulted in dark sons and blond daughters detectable by the bill and skin color at hatching.
We also had what may be termed partial sex-linked matings. These consisted of the males which were heterozygous for dark, blond or white carrying a second sex-linked color controlling gene which was recessive to that of the mother. For example, if a blond male which was heterozygous for white were mated to a blond female, then half the daughters would be white. All such whites would be female. The blond daughters could not be detected as females since the males also would be blond. (Cole____, Miller _____)
Table____
Duration: March 1974 to May
1992. Two egg clutches that have been sexed.
Hypothesis: A
1:1:1:1 ratio of these classes reflects expected equal chance
distribution of the sexes.

One expects a 1:1:1:1 ratio for these four classes. By inspection one can note the results are a good fit even without a X2 test of significance. The sex-linked results are almost too good.
Levi listed two families purporting to show a sex preference in some pigeon families. This is attributable to chance. If you have a large number of families, you can find deviations in such directions. My numbers are not so large but two samples of raw data follow.
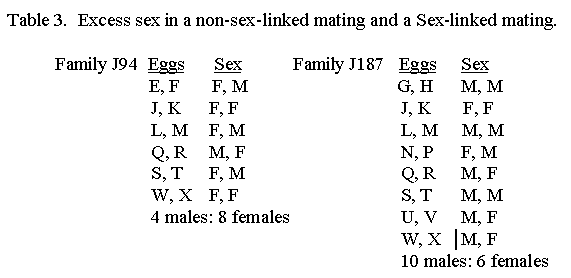
The results in the ISU colony were subject to 3 episodes of paratyphoid, Salmonella typhimurium epidemics brought in by wild house mice. This is not believed to have influenced the sex ratio. But it did reduce the numbers of 2-egg clutches successfully sexed.
A Chi Square test for the total results for J mating type birds yields a X2 of .54 with 3 degrees of freedom and a Probability of approximately .91 which is almost too good a fit.
The results for regular ringnecks, Streptopelia risoria, are summarized in --------- separating the sex-linked, partially sex-linked and autosomal results.
Duration: November 1955 to
April 1965 Families 1-300. Two egg clutches that have been
sexed.
Hypothesis: A 1:1:1:1 ratio of these classes reflects
expected equal chance distribution of the sexes.

Duration: Febuary 1968 to
July 1980 Families 308-500. Two egg clutches that have been
sexed.
Hypothesis: A 1:1:1:1 ratio of these classes reflects
expected equal chance distribution of the sexes.
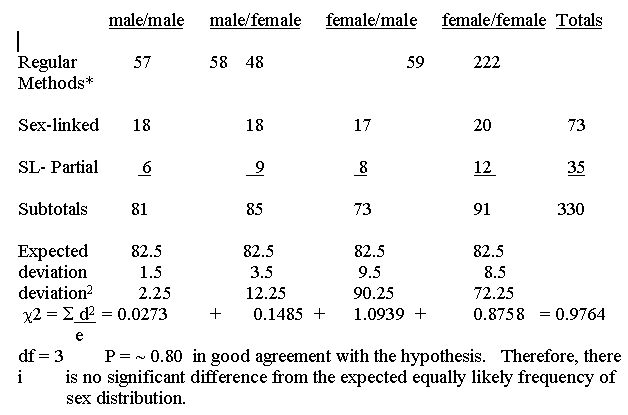
*Regular methods consist of behavior (especially bow-coos by adult males in mixed populations), vent sexing (Miller and Wagner 19 , observed copulation, actually laying eggs, production of young by pairs isolated in cages. The first two methods predominantly were used.
Duration: July 1977 to Oct 1984 ; Families 501-599 -671 June 92
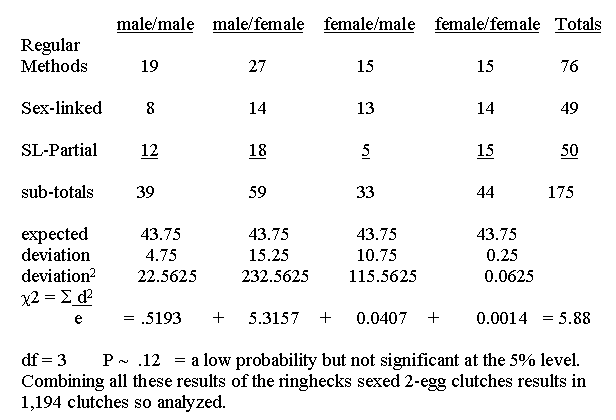
Duration:1955 to 1992
Families 1-671. Two egg clutches that have been sexed.
Hypothesis:
A 1:1:1:1 ratio of these classes reflects expected equal chance
distribution of the sexes.

Dove Behavior, Sexing Doves I, Sexing Doves II, Sexing Doves III, Sexing Doves IV, Sexing 2-egg clutches